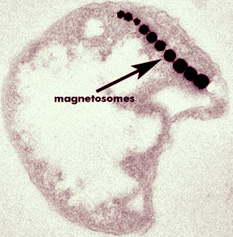
Like all magnetotactic bacteria, cells of strain MC-1 produce only one crystal morphology, in this case, elongated pseudo hexahedral prisms of magnetite. This magnetite morphology is unusual and indicates that the biomineralization processes involved in magnetosome synthesis in this bacterium are under a high degree of genetic control (Bazylinski and Frankel, 2000). There are typically 5 to 14 crystals per cell. These single-magnetic-domain crystals vary in size, averaging between 70 and 100 nm (Meldrum et al., 1993; Devouard et al., 1998). Like other magnetotactic bacteria (e.g., Magnetospirillum magnetotacticum), MC-1 cells use the chain of magnets as a compass to find favorable environmental conditions, the oxic-anoxic interface, by swimming along the Earth’s geomagnetic field lines. However, unlike cells of M. magnetotacticum which swim in both directions under oxic conditions, cells of MC-1 display a polar preference. It has been shown that magnetotaxis works in conjunction with aerotaxis (Frankel et al., 1997). Cells of M. magnetotacticum appear to use the Earth’s magnetic field lines as an axis for alignment and to rely on a temporal sensory mechanism of aerotaxis to find and maintain an optimal position in oxygen gradients; this is referred to as axial magneto-aerotaxis. Cells of MC-1 use the magnetic field directionally and appear to have a novel aerotactic sensory mechanism that seems to function as a two-way switch; this is referred to as polar magneto-aerotaxis. Cells of MC-1 also exhibit a response to short-wavelength light (< 500 nm) which causes them to swim persistently parallel to the magnetic field during illumination. Little is known of the physiology of strain MC-1. Cells of this strain are obligate microaerophiles that use thiosulfate or sulfide as electron sources (Meldrum et al., 1993) while appearing to grow autotrophically utilizing CO2 as the carbon source and producing internal sulfur globules. References: Bazylinski D. A. and Frankel R. B. (2000) Biologically controlled mineralization of magnetic iron minerals by magnetotactic bacteria. In Environmental Microbe-Metal Interactions (ed. D. R. Lovley), pp. 109-143. ASM Press. Devouard B. et al. (1998) Magnetite from magnetotactic bacteria: size distribution and twinning. Am. Mineral. 83, 1387-1398. Meldrum F. C., Mann S., Heywood B. R., Frankel R. B., and Bazylinski D. A. (1993) Electron microscopy study of magnetosomes in a cultured coccoid magnetotactic bacterium. Proc. R. Soc. Lond. B 251, 231-236. Frankel R.B., Bazylinski D. A., Johnson M. S., and Taylor B. L.. (1997) Magneto-aerotaxis in marine, coccoid bacteria. Biophys. J. 73: 994-1000. |
||
|
||
Magnetococcus MC-1