
Basidiobolus meristosporus Drechsler 1955 (synonyms
B. ranarum, B. heterosporus, B.
haptosporus) can be easy isolated from soil or leaf litter;
also from dung of frogs or lizards. It also might be found in mites
or insects, although not known to be pathogenic to all
aforementioned animals. It can be easily cultured on common
nutritious media under laboratory conditions. Basidiobolus
builds leathery, glabrous and radially folded colonies, usually
yellowish- or creamy-grey. It often forms small satellite colonies
around inoculum like sparkles due to ballistoconidia ejected from
the surface of the central colony.
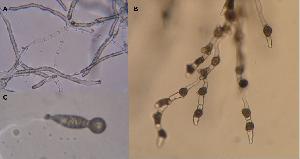
Hyphae are often septated, wide, 7-20um in diameter, many cell
compartments are without cytoplasm. Zygospores are numerous, built
between adjacent cells, 25-50um in diameter, with dark round thick
walls and beak-like appendages. Asexual spores are unicellular, of
two types: round forcibly discharged primary ballistoconidia formed
on the top of swollen conidiophores and passively released clavate
secondary conidia on non-swollen conidiophores. Ability to produce
conidia is usually lost in culture with time.
Basidiobolus has several unique features, which
distinguish it from the rest of fungal world. It has possibly the
largest nuclei among all known fungi. The structure of its mitosis
associated organelle is similar to the centrioles of all
flagellated eukaryotes. Although it is considered to be basal to
Entomophthoromycota and also has forcibly discharged conidia, the
“rocket” mechanism of the spore release has not been
found in other Entomophthoraleans.
Basidiobolus can cause infection in humans, known as
basidiobolomycoses. Infections can be subcutaneous or more rarely
gastrointestinal. Very few clinical cases have been reported,
mostly from tropical and subtropical regions.
Genome Reference(s)
Mondo SJ, Dannebaum RO, Kuo RC, Louie KB, Bewick AJ, LaButti K, Haridas S, Kuo A, Salamov A, Ahrendt SR, Lau R, Bowen BP, Lipzen A, Sullivan W, Andreopoulos BB, Clum A, Lindquist E, Daum C, Northen TR, Kunde-Ramamoorthy G, Schmitz RJ, Gryganskyi A, Culley D, Magnuson J, James TY, O'Malley MA, Stajich JE, Spatafora JW, Visel A, Grigoriev IV
Widespread adenine N6-methylation of active genes in fungi.
Nat Genet. 2017 Jun;49(6):964-968. doi: 10.1038/ng.3859